Komplexné bunkové a fyziologické procesy u živých bytostí vyžadujú jemne vyladenú reguláciu na molekulárnej úrovni, aby sa zabezpečila adaptabilita živočícha alebo rastliny, napríklad, na biotop. Na tento účel existuje mnoho molekúl, ktoré zasahujú do procesov, ako je bunková komunikácia, metabolizmus alebo delenie buniek. Jednou z týchto molekúl je proteín kalmodulinktoré pomocou vápnika ovplyvňujú funkciu mnohých ďalších biologicky aktívnych proteínov.
Čo je kalmodulín?
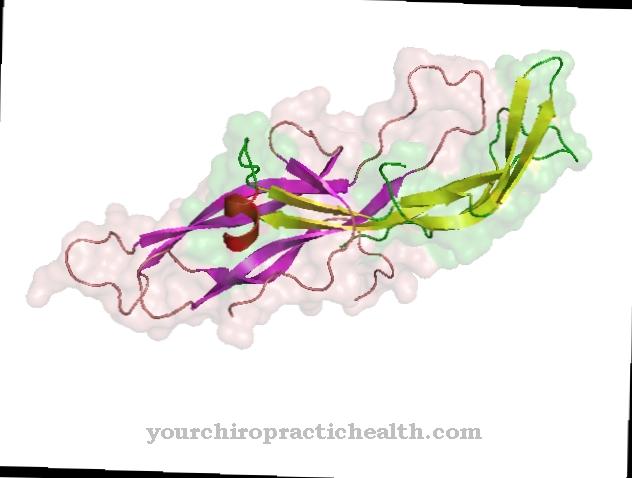
Kalmodulín je intracelulárny regulačný proteín, ktorý viaže vápenaté ióny. Vďaka svojej štruktúre patrí do skupiny proteínov EF-hand. Tvar kalmodulínu, ktorý pozostáva zo 148 aminokyselín a je dlhý 6,5 nm, sa podobá činke. Molekulová hmotnosť tejto proteínovej molekuly je asi 17 kDa.
$config[ads_text1] not found
Kalmodulín môže byť vďaka svojej biologickej funkcii pri prenose signálu v bunkách tiež klasifikovaný ako druhý messenger, t.j. sekundárna messengerová látka, ktorá však sama o sebe nie je enzymaticky aktívna. V dvoch sférických doménach proteínu sú dva motívy helix-loop-helix vo vzdialenosti 1,1 nm, na ktoré sa môžu viazať celkom štyri ióny vápnika. Táto štruktúra sa nazýva EF ruka. Štruktúry EF-hand sú spojené vodíkovými väzbami medzi antiparalelnými beta-listami kalmodulínu.
Funkcia, účinok a úlohy
Kalmodulín potrebuje tri až štyri viazané ióny vápnika na molekulu, aby bol aktívny. V aktivovanom stave sa vytvorený komplex vápnik-kalmodulín podieľa na regulácii veľkého počtu receptorov, enzýmov a iónových kanálov so širokou škálou funkcií. Medzi regulované enzýmy patrí fosfatáza kalcineurín, ktorý hrá dôležitú úlohu pri regulácii imunitnej reakcie, a endoteliálna syntáza oxidu dusnatého (eNOS), ktorá produkuje NO, ktorá sa okrem iného používa na relaxáciu hladkých svalov a tým na expanziu Cievy.
$config[ads_text2] not foundPri nízkych koncentráciách vápnika sa na druhej strane aktivuje adenylátcykláza (AC), na druhej strane, enzymatický náprotivok, fosfodiesteráza (PDE). Týmto spôsobom sa dosiahne chronologická sekvencia regulačných mechanizmov: spočiatku AC nastaví signálnu dráhu v pohybe produkciou cyklického AMP (cAMP), neskôr ju opäť vypne oponent PDE degradáciou cAMP. Regulačný účinok kalmodulínu na proteínkinázy, ako je napríklad CaM kináza II alebo kináza myozínového ľahkého reťazca (MLCK), ktorý bude podrobnejšie vysvetlený nižšie, je zvlášť známy.
CAMKII môže viazať fosfátový zvyšok na rôzne proteíny, a tým ovplyvňovať energetický metabolizmus, priepustnosť pre ióny a uvoľňovanie neurotransmiterov z buniek. CAMKII sa nachádza vo zvlášť vysokých koncentráciách v mozgu, kde hrá dôležitú úlohu v neuronálnej plasticite, t.j. všetky vzdelávacie procesy. Kalmodulín je však nevyhnutný aj pre pohybové procesy. V pokoji je koncentrácia iónov vápnika vo svalových bunkách veľmi nízka a kalmodulín je preto neaktívny. Ak je však svalová bunka vzrušená, vápnik prúdi do bunkovej plazmy a ako kofaktor zaberá štyri väzobné miesta na kalmodulíne.
To môže teraz aktivovať kinázu ľahkého reťazca myozínu, ktorá posúva sťahujúce vlákna v bunke a tým umožňuje kontrakciu svalov. Ďalšími, menej známymi enzýmami, ktoré sú ovplyvňované kalmodulínom, sú guanylátcykláza, Ca-Mg-ATPáza a fosfolipáza A2.
$config[ads_text3] not foundVzdelávanie, výskyt, vlastnosti a optimálne hodnoty
Kalmodulín sa vyskytuje vo všetkých eukaryotoch, ktoré zahŕňajú všetky rastliny, zvieratá, huby a skupinu živočíchov. Pretože molekula kalmodulínu v týchto organizmoch je zvyčajne štruktúrovaná relatívne podobným spôsobom, dá sa predpokladať, že ide o evolučne veľmi starý proteín, ktorý sa objavil v skorom štádiu.
Kalmodulín je spravidla prítomný v plazme bunky v relatívne veľkých množstvách. Napríklad v cytosóle nervových buniek je obvyklá koncentrácia okolo 30-50 uM, t.j. 0,03-0,05 mol / L. Proteín sa vytvára počas transkripcie a translácie s použitím génu CALM, ktorého doteraz sú známe tri alely, ktoré sa označujú ako CALM-1, CALM-2 a CALM-3.
Choroby a poruchy
Existujú niektoré chemikálie, ktoré môžu mať inhibičný účinok na kalmodulín, a preto sú známe ako inhibítory kalmodulínu. Ich inhibičný účinok je vo väčšine prípadov založený na skutočnosti, že transportujú vápnik z bunky a odstraňujú ho z kalmodulínu, ktorý je potom prítomný iba v neaktívnom stave.
Medzi tieto inhibujúce látky patrí napríklad W-7. Niektoré psychotropné fenotiazínové lieky navyše inhibujú kalmodulin. Pokiaľ sú regulačné funkcie kalmodulínu také široké, sú možné rôzne poruchy a poruchy, keď proteín už nie je možné aktivovať vápnikom kofaktora a regulované cieľové enzýmy sú tiež menej aktívne. Napríklad neprimeraná aktivácia CAMKII môže viesť k obmedzeniu nervovej plasticity, ktorá tvorí základ vzdelávacích procesov.
$config[ads_text4] not foundZníženie aktivácie MLCK zhoršuje kontrakciu svalov, čo môže viesť k poruchám pohybu. Menšia aktivácia enzýmu kalcineurínu v dôsledku nedostatku kalmodulínu by ovplyvnila imunitnú odpoveď tela a nižšia aktivácia eNO by viedla k nižším koncentráciám NO. Posledne menovaný spôsobuje problémy, kde má oxid dusnatý v opačnom prípade zabrániť nežiadúcemu zrážaniu krvi a rozširovať cievy za účelom lepšieho krvného obehu. V tejto chvíli by sa však malo uviesť, že kalciový senzor Frequenin môže za určitých podmienok prevziať biologické funkcie kalmodulínu, a tak nahradiť molekulu.